Specializace buněčného povrchu


Buněčný povrch se nejen v rámci specializace rozděluje na část bazální, apikální a laterální, což se týká především epitelových buněk. I buňky neepitelové však mohou mít povrchové specializace (řasinky, mikroklky apod.).
Bazální povrch
To je pol u bazální membrany. Specializací je zde bazální labyrint nebo mu taky řikají bazální žíhání. U něj buněčná membrána vbíhá do nitra buňky a tvoří četné záhyby (invaginace). Mezi záhyby jsou poté lokalizovány mitochondrie, které na tomto místě poskytují ATP pro transport iontů proti koncentračnímu gradientu, jenž je energeticky velmi náročný. V membráně labyrintu jsou umístěny i integrální membránové proteiny, které tvoří iontové pumpy. Nalezneme jej například v proximálních kanálcích ledvin.
Apikální povrch
To je pol směrem k volnému povrchu. Specializované útvary jsou zde mikroklky, stereocilie a řasinky (kinocilie).
Mikroklky

Jsou ze všech zmíněných apikálních útvarů nejmenší. Dlouhé jsou 0,5–1 μm, v průměru poté měří přibližně 0,1 μm. Jedná se o cytoplazmatické výběžky ohraničené membránou. Uvnitř obsahují dvacet až třicet aktinových mikrofilament v podélném uspořádání. Villin a fimbrin tvoří svazky filament.
Připojení mikrofilament k buněčné membráně je poté zajištěno dalším bilkovinou kalmodulinem. Pokračující ukotvení mikrofilament v rámci buňky je v terminální síti vinoucí se pod mikroklky, další opěrný bod ukotvení následně zprostředkovává aktin vázající protein fodrin, který terminální síť napojuje na intermediární filamenta.
Když velký počet mikroklků pokrývá celý apikální povrch, tak tomu řikáme kartáčový lem . Díky tomu se zvětšuje povrch a tím padem i absorbce. Najdeme ho na povrchu enterocytů v lumen střeva a u buněk vystýlajících proximální tubuly ledvin.

Stereocilie
Z výše uvedených útvarů dosahují střední délky, ta může být až 7 μm. Tyto výběžky jsou nepohyblivé, stavbou však podobné mikroklkům, tvoří je paralelně uspořádaná mikrofilamenta spojená α-aktininem. U báze jsou tlustší a následně se mohou i větvit, ve svém průběhu už ale průměr nemění. Nalezneme je v ductus epididymidis nadvarlete a ve vnitřním uchu u vláskových buněk.
Kinocilie
Kinocilie, nazývány taktéž řasinky, jsou pohyblivé cytoplazmatické výběžky tvořené složitým ciliárním aparátem. Dlouhé jsou přibližně 10 μm, v průměru dosahují tloušťky kolem 0,2 μm. Tvoří je bazální tělísko a struktury na něj navazující, což jsou: přechodná část, vlastní volná část a koncová část. K nalezení jsou (díky jejich pohyblivosti) v průdušnici, kde transportují exogenní látky z plic a ve vejcovodu, v němž napomáhají pohybu vajíčka do dělohy.

- Bazální tělísko má stejnou strukturu jako centriol. Na něj se připojují příčně pruhované kořínky směřující do cytoplazmy buňky a taktéž příčně pruhovaná bazální nožička jdoucí kolmo.
- Přechodná část řasinky je oblastí mezi bazálním tělískem a volnou částí řasinky. Nenalezneme zde centrální mikrotubuly, pouze konce mikrotubulů C tripletů stěny bazálního tělíska. Ty jsou zde uchyceny k membráně, čímž napomáhají ukotvení.

- Vlastní volná řasinka už je samotný cytoplazmatický výběžek obalený řasinkovou membránou. Střed tvoří axonema složená ze dvou centrálních mikrotubulů obklopených centrální pochvou. Okolo centrálního páru se nachází devět dvojitých periferních mikrotubulů. Periferní mikrotubuly uspořádané těsně vedle sebe jsou pokračováním mikrotubulů A a B tripletů stěny bazálního tělíska. Longitudinálně mezi nimi jsou uloženy vláknité molekuly proteinu tektinu. Mikrotubulus A je složen ze třinácti protofilament, mikrotubulus B z protofilament jedenácti, což je způsobeno tím, že část stěny má společnou s mikrotubulem A. Na mikrotubulu A jsou dvě (vnitřní a vnější) 50 nm dlouhá raménka dyneinu, motorického proteinu s vysokou ATP aktivitou. Vnější raménka jsou častější, připevňují se ke každému třetímu heterodimeru tubulinu, vnitřní raménka jsou až u každého čtvrtého heterodimeru. Spojení periferních mikrotubulů s centrální pochvou je zajištěno radiálními spojkami, ty jsou napojeny na každý čtvrtý heterodimer tubulinu. Spojení dvojitých periferních mikrotubulů navzájem je zajištěno nexinem. Ten je dlouhý 30 nm, může se však natáhnout až do délky 250 nm.
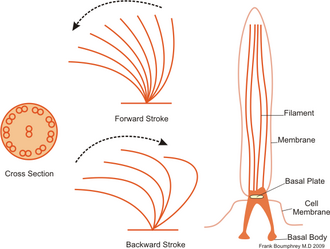
Průřez řasinkou, jejich pohyb a celková struktura řasinky.

- Koncová část řasinky je krátký úsek, kde dvojité periferní mikrotubuly přechází v jednoduché a postupně mizí. Stejně tak se zužuje i průměr řasinky. Můžeme zde někdy na povrchu plazmalemy nalézt štětičkové struktury.
Pohyb řasinek se uskutečňuje v jedné rovině, kdy rychlé ohnutí následuje postupné narovnávání. Pohyb zajišťuje vzájemný posun sousedních dvojic mikrotubulů, který vzniká dočasným spojením mezi dyneinovými raménky mikrotubulu A jedné dvojice a mikrotubulu B druhé dvojice. Vazby jsou pouze krátkodobé, rychlý vznik následuje rychlé rozpojení – tento opakující se děj lze přirovnat ke šplhání jednotlivých mikrotubulů po sobě. Motorem pohybu je MAPs dynein, jehož struktura je obdobná MAP1C dyneinu cytoplazmatickému. Jedna strana MAPs je vázána k A mikrotubulu, druhá je tvořena třemi globulárními partikulemi, které mají vysokou aktivitu ATPázy. Na ně se naváže ATP, čímž hydrolýzou dojde k uvolnění energie. Energie změní konfiguraci dyneinu, tím se protein posouvá a s ním i dvojité periferní mikrotubuly. Pohyb probíhá vždy jen v jedné polovině axonemy, tudíž je výsledný pohyb proveden pouze v jedné rovině.
Odkazy
Související články
Použitá literatura
- JUNQUIERA, L. Carlos, José CARNEIRO a Robert O. KELLEY. Základy histologie. 1. vydání. Jinočany : H & H 1997, 1997. 502 s. ISBN 80-85787-37-7.
- LÜLLMANN-RAUCH, Renate. Histologie. 1. vydání. Praha : Grada, 2012. 576 s. ISBN 978-80-247-3729-4.
